Ученые из Московского физико-технического института, Института биоорганической химии им. М. М. Шемякина и Ю. А. Овчинникова и НИИ системной биологии и медицины с коллегами из Южной Кореи и США выяснили, как холестерин на цитоплазматической стороне клеточной мембраны управляет работой рецептора эпидермального фактора роста (EGFR). Этот рецептор отвечает за деление клеток, а его избыточная активность связана с развитием онкологических заболеваний.
Оказалось, что холестерин специфически встраивается в специальный «карман» между мембранными сегментами рецептора, стабилизируя рецептор в активной конформации для проведения биохимического сигнала внутрь клетки. Исследование выполнено при поддержке РНФ (грант № 23‑74‑00024), результаты опубликованы в журнале Science Advances.
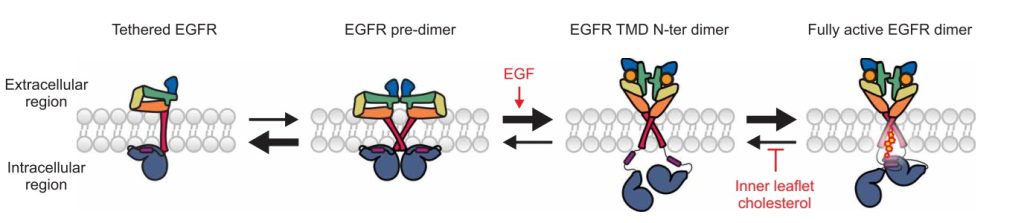
Рецептор эпидермального фактора роста (EGFR) — трансмембранный белок, который отвечает за «чувствительность» клетки к молекулярным биохимическим сигналам, запускающим клеточное деление. Нарушение работы рецептора, его чрезмерная активность приводит к тому, что клетка бесконтрольно размножается, а это может привести к образованию раковой опухоли. Сигнал к работе подает молекула эпидермального фактора роста (EGF), но одного сигнала недостаточно — рецептор должен «сложиться» в определенную конформацию.
Биофизики из Центра исследований молекулярных механизмов старения и возрастных заболеваний МФТИ, Института биоорганической химии им. М. М. Шемякина и Ю. А. Овчинникова и НИИ системной биологии с коллегами из Южной Кореи выяснили, что важнейшую роль в этом процессе играет холестерин на внутренней цитоплазматической стороне клеточной мембраны.
Ранее считалось, что холестерин в мембране только ингибирует активность EGFR, который до встречи с лигандом преимущественно располагается в мембранных рафтах, насыщенных холестерином, и часто существует в виде неактивного пред-димера.
В исследовании ученые убедительно доказали, что та же молекула холестерина с внутренней цитоплазматической стороны мембраны служит своеобразным «замковым камнем», стабилизирующим уже активную димерную конформацию EGFR.
Как только димер EGFR связывается с фактором роста EGF, внеклеточные домены рецептора меняют конформацию, что инициирует проворот трансмембранных спиралей относительно друг друга и дальнейшее их переключение в активную димерную конформацию с формированием специфического гидрофобного «кармана» в цитоплазматической части мембраны. Именно туда садится одна молекула холестерина, прочно фиксируя рабочее состояние рецептора.
Без такой поддержки активный димер EGFR остается нестабильным, что, в свою очередь, не позволяет примембранным участкам и киназным доменам рецептора уложится в определенную структуру, необходимую для аутофосфорилирования и проведения внутриклеточного сигнала, управляющего делением клетки.
Авторы работы адаптировали созданный ими ранее метод ко-иммобилизации одиночных молекул (Co‑II) для измерения белок-белковых контактов, заменив один из белков флуоресцентно меченым липидом, что впервые позволило в реальном времени количественно оценить взаимодействие между отдельной молекулой холестерина и рецептором прямо в плазматической мембране живой клетки.
Суть метода состоит в том, что на стеклянной поверхности, на которой лежат живые клетки, закреплены вторичные антитела. Когда в среду добавляют первичные антитела против определенной метки на «приманочном» (bait) белке, все молекулы этого белка оказываются буквально пришитыми к стеклу через цепочку антител. Если липидные молекулы-«жертвы» (prey), меченные флуоресцентной меткой, взаимодействуют с иммобилизованным белком-приманкой, они тоже становятся неподвижными. Таким образом, Co‑II дает возможность буквально «увидеть» момент связывания и подсчитать, какая доля молекул липида вовлечена в комплекс.
В качестве белка-приманки ученые использовали рецептор фактора роста, к которому с помощью методов генной инженерии был «пришит» белок SNAP. SNAP-метка — это «клей» на основе человеческого белка (O6-алкилгуанин-ДНК-алкилтрансферазы), которая позволяет надежно присоединять к белку-носителю любую синтетическую молекулу (например, флуоресцентный краситель CF660R), к которой химически привязан бензилгуанин. К «жертве»-холестерину присоединили флуоресцентный краситель BODIPY, и таким образом, за перемещением одиночных молекул BODIPY‑холестерина и SNAP‑EGFR стало возможно наблюдать с помощью конфокального флуоресцентного микроскопа.
Прослеживая историю каждой молекулы, авторы могли точно разделить подвижные и неподвижные молекулярные комплексы. Сначала в отсутствие иммобилизующих антител «жертвы» и «приманки» свободно перемещались по мембране. Затем ученые «приклеили» модифицированный рецептор фактора роста к стеклу с помощью антител к SNAP-метке, повторно пронаблюдали траектории движения BODIPY‑холестерина в той же самой клетке и вычислили долю холестериновых частиц, которые стали неподвижными именно из-за связывания с рецептором. Оказалось, что в отсутствие фактора роста EGF эта доля неотличима от нуля, а после добавления EGF около 6–7 % BODIPY‑холестерина иммобилизуется вместе с активным EGFR.
Но что, если меченый холестерин взаимодействует с рецептором не так, как природный? Чтобы выяснить это, ученые сперва удалили весь холестерин из плазматической мембраны с помощью метил‑β‑циклодекстрина (MβCD). Это спровоцировало лиганд‑независимую, спонтанную активацию рецептора — EGFR начинал сигналить на некотором уровне даже без эпидермального фактора роста.
Затем в мембрану «возвращали» либо обычный холестерин, либо BODIPY‑меченый, и смотрели, подавляется ли эта паразитная активность. Оказалось, что оба варианта холестерина одинаково эффективно гасили несанкционированную активацию рецептора. Это значит, что BODIPY‑Chol правильно встраивается в липидный бислой, корректно взаимодействует с EGFR и выполняет ту же регуляторную роль, что и нативный холестерин.
Далее исследователи задались вопросом, на каком этапе холестерин взаимодействует с рецептором? Ранее проведенные исследования свидетельствовали о том, что факт взаимодействия проявлялся только после добавления эпидермального фактора роста.
Классическая модель работы EGFR гласит, что EGF запускает димеризацию рецептора и переводят его в активное состояние. Если холестерин начинал «прилипать» к EGFR лишь после стимуляции лигандом, логично было предположить, что мишенью служит именно активный димер, а не покоящийся мономер рецептора. Чтобы убедиться в этом, исследователи «подсунули» холестерину мутантную форму EGFR.
У такого рецептора внеклеточная часть всегда находится в вытянутой, «расправленной» конформации, которая характерна для активированного димера, и это состояние не требует добавления фактора роста. Оказалось, что холестерин связывался с этим мутантом даже без добавления EGF.
То же самое показали для ErbB2 — близкого родственника EGFR (ErbB1), чей внеклеточный домен от природы существует в вытянутой, активноподобной форме, и который склонен к димеризации в активную конформацию. Раз холестерин взаимодействовал с заведомо димеризованными или предрасположенными к димеризации формами, значит, ему нужен именно димер.
Окончательно устранить сомнения помогли опыты с фотообесцвечиванием. Исследователи использовали дестиобиотин (химически модифицированная форма биотина, витамина H), чтобы обратимо прикреплять рецепторы к стеклу и аккуратно снижать их плотность до уровня, при котором в объективе микроскопа видны отдельные молекулы. Затем они следили за тем, как флуоресцентная метка на SNAP-EGFR постепенно выцветает (фотообесцвечивается) под лазером.
Если в светящейся точке находится одиночная молекула белка, ее сигнал исчезает резко, за один шаг. Если же точка содержит димер — две молекулы EGFR — флуоресценция падает в два последовательных шага, когда каждая из молекул по очереди теряет свечение.
Подсчитав долю «одношаговых» и «двухшаговых» пятен, авторы определили, что без эпидермального фактора роста (EGF) около 72,5% рецепторов находятся в мономерном состоянии, а около 21,5% — в виде неактивных пред-димеров. После добавления EGF доля димеров возрастает более чем вдвое — до 45,9%. Это прямое доказательство того, что активация EGFR вызывает его димеризацию, и что холестерин взаимодействует именно с этой активной димерной формой рецептора.
Дальше в дело вступили компьютерное моделирование и ядерный магнитный резонанс (ЯМР). С помощью молекулярной динамики исследователи построили модель активной формы димера, и увидели, что между расправленными трансмембранными спиралями образуется полость, куда заходит одна молекула холестерина и остается там.
Холестерин контактирует с тремя аминокислотными остатками — изолейцином-640, глицином-641 и лейцином-642, это порядковые номера конкретных «кирпичиков» в структуре рецептора, за которые цепляется холестерин. Благодаря этому трансмембранные спирали слегка раздвигаются своими нижними концами, и части рецептора, содержащие киназные домены, которые находятся внутри клетки и отвечают за передачу сигнала, получают возможность сблизиться в определенной конформации. Это запускает химическую реакцию присоединения фосфатных групп (фосфорилирование) — главный признак того, что рецептор включился и дал клетке команду делиться.
Чтобы проверить, действительно ли эти три аминокислоты отвечают за связывание холестерина, ученые сделали точечные мутации в гене EGFR. Замена изолейцина-640, глицина-641 или лейцина-642 на другие аминокислоты приводила к тому, что рецептор переставал связывать холестерин и не активировался даже в присутствии фактора роста.
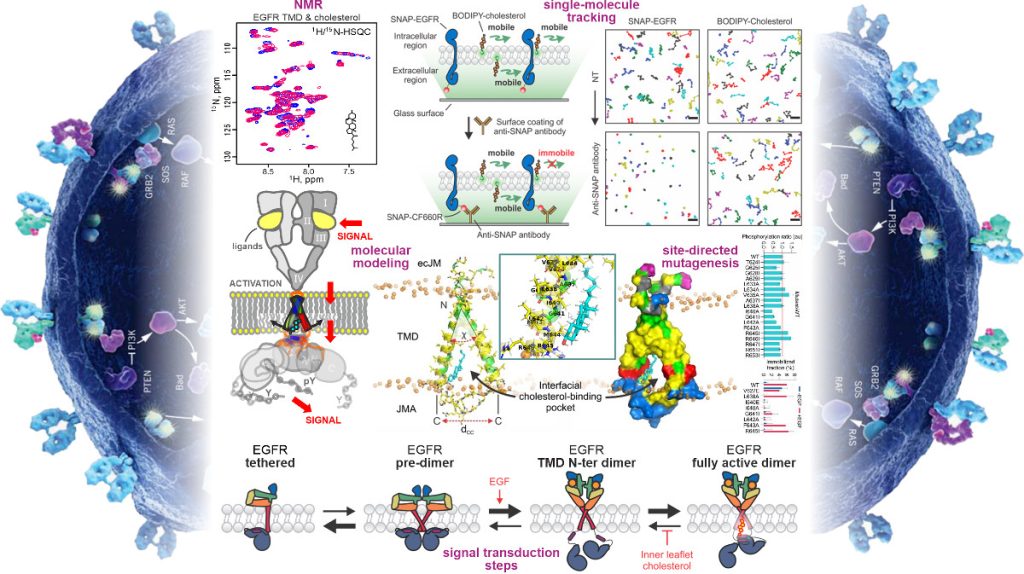
2) Метод визуализации в живых клетках: рецептор EGFR (меченный SNAP-меткой) и холестерин (меченный зеленым флуорофором BODIPY) отслеживаются в реальном времени. Клеточная мембрана изображена в виде двух слоев, а отдельные точки — это траектории движения единичных молекул. Метод показывает, что холестерин связывается только с активной формой EGFR.
3) Сигнальный каскад: Упрощенная схема передачи сигнала в клетку. Активация EGFR (желтая молния) запускает цепочку взаимодействий белков (GRB2, SOS, RAF), что в конечном итоге приводит к включению генов, контролирующих рост и выживание клетки.
4) Молекулярная модель связывания: Трехмерная структура трансмембранного домена димера EGFR (две «подковы») в мембране (фосфатные группы липидов показаны желтыми шариками). После связывания EGF рецептором молекула холестерина (выделена голубым) заходит в сформированную полость между мембранными сегментами рецептора, действуя как «молекулярный замок», стабилизирующий его активную конформацию. Красным выделены критические аминокислоты (Ile640-Gly641-Leu642), мутации в которых разрушают связывание холестерина и блокируют активацию EGFR. Источник: Science Advances
«Мы показали, что холестерин во внутренней цитоплазматической части мембраны — не просто пассивная добавка, а полноценный регуляторный переключатель,— пояснил Эдуард Бочаров, старший научный сотрудник Центра исследований молекулярных механизмов старения и возрастных заболеваний МФТИ, заведующий лабораторией биомолекулярной ЯМР-спектроскопии ГНЦ ИБХ РАН.
— Без него активная конформация димера рецептора EGFR нестабильна, что не позволяет надежно провести через мембрану биохимический сигнал на пролиферацию клетки. Это указывает на согласованность белок-липидных взаимодействий и раскрывает биофизические принципы передачи сигнала через клеточную мембрану. Кроме того, данный факт дает молекулярное обоснование тому, почему статины — лекарства, снижающие холестерин — могут подавлять рост некоторых раковых клеток. Вероятно, их терапевтический эффект частично связан с дефицитом холестерина именно во внутреннем слое мембраны, что мешает рецепторам EGFR полноценно «включаться»».
Полученные результаты открывают новую мишень для противораковой терапии — карман между трансмембранными доменами EGFR, куда садится холестерин. Если найти молекулу, которая модифицирует этот карман, можно попытаться заблокировать рецептор, который уже активирован сигналом. Также в ближайших планах исследователей — проверить, работает ли такой же механизм у других рецепторов семейства ErbB, например, у ErbB2 (HER2), который является онкомаркером и вовлечен в патогенез рака груди.
Исследование выполнено при поддержке РНФ (грант № 23‑74‑00024).
Научная статья: Do-Hyeon Kim, Minh-Triet Hong, Eduard V. Bocharov, Soyeon Park, Yaroslav V. Bershatsky, Pavel E. Volynsky, Eui Kim, Roman G. Efremov, Wonhwa Cho; EGFR activation requires cholesterol interaction at the inner leaflet of the plasma membrane. Sci. Adv.11, (2025).
DOI: 10.1126/sciadv.adx2398